
.jpg)
Hotline
024.3212.3709
Kiểm định nguồn gốc dược liệu đang được coi là một vấn đề nóng hiện nay. Trên thế giới, các nhà khoa học đang rất nỗ lực trong việc kiểm định các giống sâm nhằm phân loại một cách chính xác, thuận tiện và nhanh chóng nguồn gốc của chúng. Từ những kết quả đạt được trong nghiên cứu giám định các loài sâm phổ biến trên thế giới, các tác giả đã đề xuất quy trình kỹ thuật đạt chuẩn quốc tế cho việc giám định sâm Ngọc Linh ở Việt Nam. Đây là những bước đi rất có ý nghĩa nhằm nâng cao giá trị thương hiệu sâm Ngọc Linh của nước ta trên thị trường trong nước và quốc tế.
Mở đầu
Trên thế giới, nhu cầu sử dụng thuốc bào chế từ dược liệu rất lớn, đó là chưa kể dược liệu dùng trong sinh hoạt hàng ngày với tác dụng bồi bổ cơ thể. Đây rõ ràng là lợi thế và cơ hội rất lớn cho ngành dược liệu Việt Nam. Cây dược liệu có thể được trồng và sinh trưởng tốt ở những khu vực núi cao, nơi khó canh tác nông nghiệp một cách hiệu quả. Vì thế, chúng hoàn toàn có thể mang lại nguồn thu lớn, trở thành cây xóa đói giảm nghèo cho nông dân nếu được định hướng và chỉ đạo rõ ràng. Tuy nhiên, một trong những vấn đề bức xúc và đáng lo ngại là sự thật - giả về nguồn gốc dược liệu quý trên thị trường cung ứng hiện nay, đặc biệt đối với cây sâm Ngọc Linh (Panax vietnamensis Ha et Grushv).
Để tìm hiểu về vấn đề kiểm định sự thật - giả của dược liệu, bên cạnh phương pháp nhận dạng hình thái, xác định hoạt chất, một trong những điểm mấu chốt là đặc điểm di truyền đặc trưng của từng loại cây dược liệu. Cho đến nay, rất nhiều phương pháp đã được áp dụng để nhận biết những điểm đặc thù trên phân tử ADN di truyền từ thế hệ này sang thế hệ khác của loài.
Một số đặc điểm di truyền hệ gen lục lạp của các loài sâm
Nằm trong bản đồ phân bố sâm của thế giới, Việt Nam với điều kiện tự nhiên đa dạng là nơi phân bố của một số loài sâm có giá trị cao như: Vũ Diệp (P. bipinnatifidus Seem.) ở Hoàng Liên Sơn, Tam thất (P. pseudoginseng Wall.) ở Sa Pa, Nhật (P. japonicus) và Ngọc Linh ở Kon Tum và Quảng Nam. Rõ ràng, việc trà trộn sâm Ngọc Linh với những loại sâm nêu trên, thậm chí với một số loại củ không thuộc chi Panax là điều hoàn toàn có thể xảy ra trong khi chất lượng và hoạt tính giữa chúng rất khác biệt.
Cơ sở khoa học của giám định gen chủ yếu dựa trên hệ gen nhân và lục lạp của các đối tượng nghiên cứu. Vì thế, hệ gen lục lạp của một số loài sâm cũng đã bắt đầu được giải trình tự một cách đầy đủ (hình 1A). Năm 2004, hệ gen lục lạp của sâm Hàn Quốc (P. schinseng Nees) có kích thước sợi ADN mạch vòng đạt 156.318 bp đã được ghi nhận đầu tiên [1]. Sau đó, 4 nòi sinh thái của P. ginseng (Damaya, Ermaya, Gaolishen và Yeshanshen) cũng lần lượt được giải trình tự hệ gen lục lạp để dự đoán về cơ chế tiến hóa và mức độ đa hình giữa chúng [2]. Cụ thể, Damaya, Ermaya và Gaolishen có kích thước hệ gen lục lạp là 156.354 bp, trong khi kích thước sợi ADN mạch vòng ở nòi Yeshanshen là 156.355 bp [2]. Đến nay, sâm Mỹ (P. quinquefolius, P. notoginseng, P. japonicus) cũng đã được tiến hành giải trình tự hệ gen lục lạp, có kích thước lần lượt là 156.088, 156.466 và 156.188 bp [3-6]. Đáng chú ý, sâm Ngọc Linh của Việt Nam cũng đã được giải trình tự hệ gen lục lạp một cách hoàn chỉnh [5, 6]. Kích thước sợi ADN mạch vòng có chiều dài 155.993 bp, chứa 79 gen mã hóa protein, 29 gen mã hóa phân tử tARN và 4 gen mã hóa rARN [6]. Kết quả xây dựng cây phân loại cho thấy, sâm Ngọc Linh có nguồn gốc tiến hóa gần gũi với P. notoginseng và P. japonicus (hình 1B). Những kết quả này có thể mở ra những bí mật về cơ chế quang hợp của họ Panax và quan trọng hơn cả là cho phép phân biệt chúng ở cấp độ phân tử dựa vào sự sai khác giữa các loài.
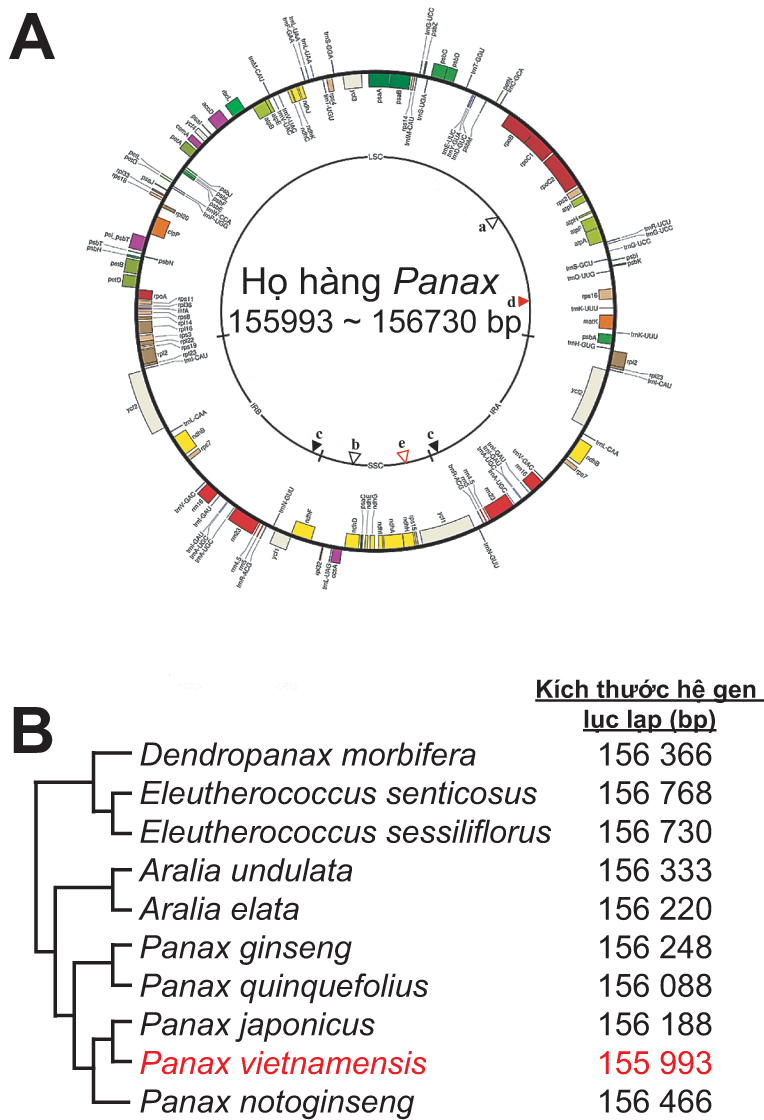
Gần đây, một số loài cây thuốc trong chi có họ hàng gần gũi với Panax spp., như Eleutherococcus, Aralia và Dendropanax cũng được quan tâm và nghiên cứu để cung cấp thêm một cách rõ nét về sự đa dạng di truyền và phân loại giữa các họ hàng của Panax ở cấp độ phân tử [5]. Đi sâu hơn vào việc nhận biết những vùng bảo thủ đặc trưng giữa các loài, Kim và cộng sự cũng đã tìm ra được rất nhiều vị trí đa hình ở các gen mã hóa nrARN 45S (nuclear ribosomal ARN 45S) giữa 10 loài thuộc Panax spp. và 3 chi họ hàng. Những ghi nhận bước đầu này đã đặt nền móng quan trọng cho việc kiểm định sự có mặt của sâm Ngọc Linh với những loài gần gũi với chi Panax trong sản phẩm. Từ đây, một số thành tựu trong nghiên cứu giám định sâm đã được báo cáo trên thế giới.
Một số kết quả trong nghiên cứu giám định sâm trên thế giới
Giám định sâm, trước hết phải dựa vào đặc điểm hình thái đặc trưng của từng loài hoặc phương pháp phân tích protein để xác định sự có mặt của hợp chất quý trong sâm. Bên cạnh đó, phương pháp xác định nhờ chỉ thị phân tử ADN đã được sử dụng rất phổ biến ở đối tượng cây trồng để tìm hiểu bản chất di truyền và xác định các locus gen quy định tính trạng. Các loại chỉ thị phân tử được sử dụng phổ biến trong nhận dạng sâm bao gồm RFLP (Restriction fragment length polymorphism), RAPD (Random amplified polymorphic DNA), STS (Sequence - tagged site), SSR (Simple sequence repeats) và SNP (Single nucleotide polymorphisms). Một số nghiên cứu đã thiết kế chỉ thị RFLP để nhận diện sự sai khác giữa một số vùng đặc trưng, như gen 18S rARN, trình tự ribosome ITS1-5.8S-ITS2, trình tự rARN 5S [7]. Bên cạnh đó, chỉ thị SSR được sử dụng phổ biến hơn cả trong phân tích đa dạng di truyền giữa các giống sâm do đây là các đoạn trình tự lặp đơn, ngắn nên rất đặc hiệu cho từng giống [7, 8]. Liên quan đến vấn đề bảo hộ sản phẩm sâm Hàn Quốc, Kim và cộng sự cũng đã phát triển 19 chỉ thị EST-SSR để kiểm định 9 giống sâm trồng tại Hàn Quốc [9]. Đây được xem là tiền đề quan trọng cho công tác chọn tạo giống sâm chất lượng cao tại Hàn Quốc. Gần đây, với sự phát triển của công nghệ giải trình tự thế hệ mới đã cho phép xác định những điểm sai khác giữa các giống sâm, các sai khác này có thể được rà soát và xác định bằng chỉ thị SNP [7, 10]. Trong bối cảnh mở cửa thị trường trao đổi hàng hóa tự do, ngành công nghiệp sâm ở các nước đã bị tác động không nhỏ từ việc trộn lẫn và làm giả các loại sâm. Vì vậy, cần thiết phải sàng lọc ra các chỉ thị phân tử ADN để nhận diện giữa các giống cũng như xây dựng cơ sở dữ liệu phân tử của các loài. So với tốc độ phát triển của ngành công nghiệp sâm, phương pháp phân tích ADN truyền thống được cho là tốn kém, cần nhiều thời gian trong khi hiệu quả lại không cao. Gần đây, công nghệ giải trình tự thế hệ mới đã cho phép chúng ta dễ dàng xác định một cách nhanh chóng các đột biến như SNP, SSR, thêm/mất ở trình tự hệ gen.
Tiếp theo, một kỹ thuật được sử dụng khá phổ biến trong nhận dạng sâm là barcode (250-1000 bp) và mini-barcode (100-250 bp). Một số kết quả bước đầu đã được ghi nhận trong nỗ lực định danh các loài sâm khác nhau, bao gồm 33 nòi sinh thái P. bipinnatifidus, 2 loài sâm P. ginseng, P. notoginseng thu thập tại Trung Quốc, 1 loài P. pseudoginseng thu thập tại Nepal, sâm Mỹ P. quinquefolius và P. trifolius, 5 nòi sinh thái P. japonicus và sâm P. stipuleanatus. Đáng chú ý, có 3 mẫu sâm được thu thập tại Việt Nam, là P. stipuleanatus (Quảng Nam) và 2 mẫu P. bipinnatifidus ghi nhận ở Lâm Đồng và Quảng Nam [11]. Gần đây, mini-barcode cũng đã được phát triển để nhận dạng P. notoginseng [3].
Mới đây, nhóm nghiên cứu tại Đại học Quốc gia Seoul (Hàn Quốc) đã công bố thành công trong việc giám định 5 loại sâm P. ginseng, P. quinquefolius, P. notoginseng, P. japonicus và Ngọc Linh [12]. Trong đó, 14 chỉ thị InDel cho kết quả đa hình giữa 5 hệ gen lục lạp đã được xác định để giám định Panax spp. Đáng chú ý, 2 chỉ thị phân tử, bao gồm gcpm9 (thiết kế từ đoạn 25 bp TR và 6 bp SSR trên vùng clpP-psbB) và gcpm14 (thiết kế từ đoạn 30 bp InDel trên vùng ycf1) được xác định rất đặc trưng cho sâm Ngọc Linh [12]. Để tránh sự trộn lẫn thành phần với các loài khác trong họ Araliaceae, nhóm nghiên cứu đã tiếp tục đánh giá sự sai khác giữa hệ gen lục lạp và nrARN giữa các loài có họ hàng gần gũi Panax spp., Eleutherococcus spp., Aralia spp. và Dendropanax spp. [5].
Cuối cùng, khi câu chuyện về giám định các giống sâm đã gần như sáng tỏ, việc mã hóa các kết quả thu được dưới dạng mã phản hồi nhanh (QR code) tiếp tục được quan tâm. Một nghiên cứu gần đây đã công bố về việc chuyển đổi kết quả giám định giống P. ginseng thành dạng QR code bằng các thuật toán 2 chiều [13]. Đây được xem là tiền đề rất quan trọng để người tiêu dùng có thể tự đánh giá và kiểm định chất lượng của sản phẩm sâm trên thị trường thông qua QR code.
Đề xuất quy trình giám định sâm Ngọc Linh ở Việt Nam
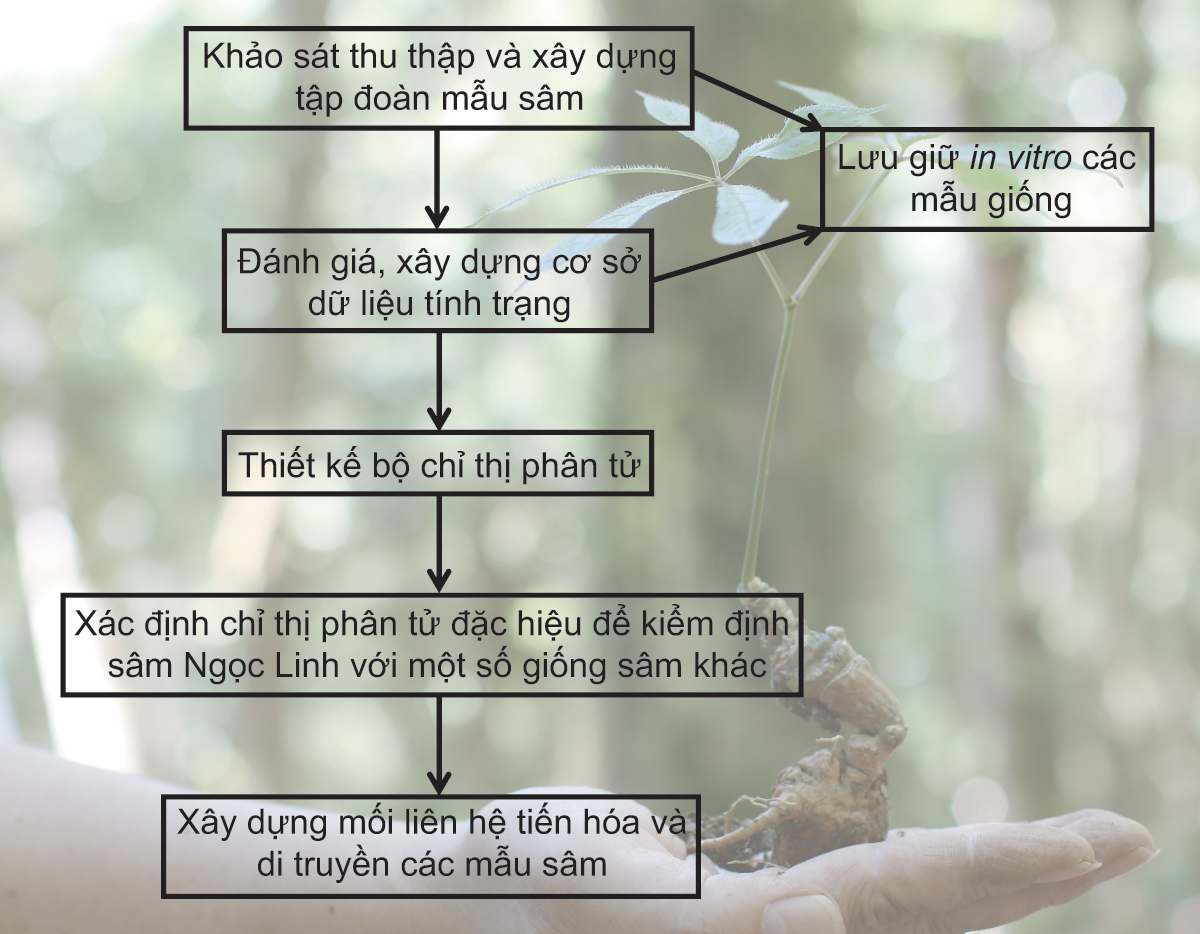
Để ngăn chặn sự thất thoát nguồn sâm tự nhiên và hiện tượng trà trộn làm giả các sản phẩm sâm Ngọc Linh, cần khẩn trương xây dựng tập đoàn giống gốc cây sâm Ngọc Linh có sức chống chịu tốt, sinh trưởng nhanh, năng suất cao phục vụ công tác bảo tồn và nhân giống. Câu hỏi đang được quan tâm hiện nay là làm thế nào để phân biệt sâm Ngọc Linh thật và các loài sâm Panax spp. ở Việt Nam? Để trả lời được câu hỏi này, bộ chỉ thị phân tử ADN dựa trên trình tự hệ gen sẽ được sử dụng trong nghiên cứu di truyền phân tử và xác định chỉ thị phân tử đặc hiệu nhằm kiểm định sâm Ngọc Linh phục vụ nhu cầu sản xuất và bảo đảm quyền lợi người tiêu dùng. Đây là những nội dung chính cần phải đạt được để quản lý và khai thác sâm Ngọc Linh một cách hợp lý và bền vững (hình 2). Cụ thể là:
Một là, thu thập và xây dựng tập đoàn mẫu sâm Ngọc Linh, Vũ Diệp, Tam thất hoang Lai Châu và sâm Hàn Quốc. Khảo sát đa dạng nguồn sâm phân bố tại các khu vực sinh thái khác nhau trên những vùng núi cao, từ đó nắm được thực trạng nguồn sâm phân bố ở Việt Nam hiện nay.
Hai là, nghiên cứu đánh giá các tính trạng hình thái sinh học chính, từ đó xây dựng hệ thống cơ sở dữ liệu của các mẫu sâm Ngọc Linh thu thập được ở các vùng sinh thái khác nhau. Việc thiết lập và xây dựng dữ liệu kiểu hình các tính trạng của mẫu sâm Ngọc Linh là tiền đề quan trọng cho công tác nhận dạng và phát triển giống trong tương lai.
Ba là, nghiên cứu lưu giữ in vitro các mẫu sâm thu thập được. Song song với việc thu thập, công tác lưu giữ và phục tráng bằng các kỹ thuật nuôi cấy mô tế bào thực vật cần được tiến hành nhằm bảo quản toàn bộ ngân hàng mẫu giống, phục vụ chọn tạo và phát triển sau này. Việc tối ưu hóa môi trường dinh dưỡng và điều kiện nuôi cấy cũng được quan tâm nhằm tăng hiệu quả và khả năng phát sinh chồi từ các mẫu sâm thu thập được, từ đó hoàn thiện quy trình tạo cây in vitro.
Bốn là, thiết kế bộ chỉ thị phân tử phục vụ nghiên cứu di truyền và kiểm định sâm Ngọc Linh. Các chỉ thị phân tử được xác định và thiết kế dựa trên hệ gen nhân và hệ gen lục lạp của sâm Ngọc Linh. Sau đó, các chỉ thị đặc hiệu được chọn lọc để phân biệt sâm Ngọc Linh với một số giống sâm khác. Từ những kết quả thu được, bản đồ di truyền liên kết với các tính trạng quan trọng sẽ được thiết lập, từ đó phục vụ công tác lai tạo giống sâm chất lượng cao.
Đây là những nội dung chính để xây dựng một quy trình kỹ thuật đạt chuẩn quốc tế cho việc kiểm định giống sâm Ngọc Linh ở mức độ phân tử. Những kết quả thu được sẽ đóng góp rất có ý nghĩa trong việc nâng cao giá trị của sâm Ngọc Linh.
TÀI LIỆU THAM KHẢO
[1] K.J. Kim, H.L. Lee (2004), “Complete chloroplast genome sequences from Korean ginseng (Panax schinseng Nees) and comparative analysis of sequence evolution among 17 vascular plants”, DNA Res., 11(4), pp.247-261.
[2] Y. Zhao, J. Yin, H. Guo, Y. Zhang, W. Xiao, C. Sun, J. Wu, X. Qu, J. Yu, X. Wang, J. Xiao (2014), “The complete chloroplast genome provides insight into the evolution and polymorphism of Panax ginseng”, Front. Plant Sci., 5, doi: 10.3389/fpls.2014.00696.
[3] W. Dong, H. Liu, C. Xu, Y. Zuo, Z. Chen, S. Zhou (2014), “A chloroplast genomic strategy for designing taxon specific DNA mini-barcodes: A case study on ginsengs”, BMC Genet., 15, doi.org/10.1186/s12863-014-0138-z.
[4] Z.J. Han, W. Li, Y. Liu, L.Z. Gao (2015), “The complete chloroplast genome of North American ginseng, Panax quinquefolius”, Mitochondrial DNA, 27, pp.3496-3497.
[5] K. Kim, V.B. Nguyen, J. Dong, Y. Wang, J.Y. Park, S.C. Lee, T.J. Yang (2017), “Evolution of the Araliaceae family inferred from complete chloroplast genomes and 45S nrDNAs of 10 Panax - related species”, Sci. Rep., 7, pp.1-9.
[6] B. Nguyen, K. Kim, Y.C. Kim, S.C. Lee, J.E. Shin, J. Lee, N.H. Kim, W. Jang, H.I. Choi, T.J. Yang (2015), “The complete chloroplast genome sequence of Panax vietnamensis Ha et Grushv (Araliaceae)”, Mitochondrial DNA, 28, pp.1-2.
[7] I.H. Jo, Y.C. Kim, D.H. Kim, K.H. Kim, T.K. Hyun, H. Ryu, K.H. Bang (2016), “Applications of molecular markers in the discrimination of Panax species and Korean ginseng cultivars (Panax ginseng)”, J. Ginseng Res., 41(4), pp.444-449.
[8] H.I. Choi, N.H. Kim, J.H. Kim, B.S. Choi, I.O. Ahn, J.S. Lee, T.J. Yang (2011), “Development of reproducible EST-derived SSR markers and assessment of genetic diversity in Panax ginseng cultivars and related species”, J. Ginseng Res., 35, pp.399-412.
[9] N.H. Kim, H.I. Choi, I.O. Ahn, T.J. Yang (2012), “EST-SSR marker sets for practical authentication of all nine registered ginseng cultivars in Korea”, J. Ginseng Res., 36, pp.298-307.
[10] Y. Liu, X. Wang, L. Wang, X. Chen, X. Pang, J. Han (2016), “A nucleotide signature for the identification of American ginseng and its products”, Front Plant Sci, 7, doi. 10.3389/fpls.2016.00319.
[11] Y.J. Zuo, Z.J. Chen, K. Kondo, T. Funamoto, J. Wen, S.L. Zhou (2011), “DNA barcoding of Panax species”, Planta Med., 77, pp.182-187.
[12] V.B. Nguyen, H.S. Park, S.C. Lee, J. Lee, J.Y. Park, T.J. Yang (2017), “Authentication markers for five major Panax species developed via comparative analysis of complete chloroplast genome sequences”, J. Agric. Food Chem., 65, pp.6298-6306.
[13] Y. Cai, P. Li, X.W. Li, J. Zhao, H. Chen, Q. Yang, H. Hu (2017), “Converting Panax ginseng DNA and chemical fingerprints into two-dimensional barcode”, J. Ginseng Res., 41, pp.339-346.
- Chu Đức Hà và cộng sự -

Sáng nay, Khoa Công nghệ Nông nghiệp trang trọng tổ chức Lễ bế giảng khóa đào tạo ngắn hạn về kỹ năng phân tích RNA-Seq, chương trình phối hợp cùng Công ty LOBI Việt Nam – doanh nghiệp tiên phong trong phát triển giải pháp tin sinh học.

Buổi seminar khoa học của Khoa Công nghệ Nông nghiệp đã diễn ra thành công tốt đẹp với phần trình bày chuyên sâu từ Dr. Pierre Larmande – Senior Research Scientist, Viện Nghiên cứu Phát triển Bền vững Pháp (IRD).

Khoa Công nghệ Nông nghiệp trân trọng kính mời quý thầy cô và toàn thể sinh viên các khóa, đặc biệt là sinh viên định hướng Tin sinh học (Bioinformatics) tham dự buổi báo cáo khoa học do Dr. Pierre Larmande – Senior Research Scientist tại Viện Nghiên cứu Phát triển Bền vững Pháp (IRD) trình bày.

Nhóm nghiên cứu của Khoa Công nghệ Nông nghiệp đã có phần báo cáo tại Hội thảo Khoa học Khoa Sinh học 2025 của Trường Đại học Sư phạm Hà Nội 2, với chủ đề “Nghiên cứu và giảng dạy Khoa học sự sống trong thời đại 4.0”.

Nhóm nghiên cứu Khoa Công nghệ Nông nghiệp – Trường Đại học Công nghệ (ĐHQGHN) vừa có buổi làm việc với Bộ môn Công nghệ Sinh học – Trường Đại học Thủy lợi.

Trong hai ngày 17–18/10/2025, tại Trường Đại học Nông Lâm TP. Hồ Chí Minh, Hội nghị Công nghệ Sinh học toàn quốc năm 2025 đã diễn ra sôi nổi, quy tụ hàng trăm nhà khoa học, chuyên gia và giảng viên đến từ các trường đại học, viện nghiên cứu trên cả nước.


Khoa Công nghệ Nông nghiệp chính thức ra mắt Câu lạc bộ sinh viên PYLIFE – không gian dành cho những bạn trẻ đam mê lập trình ứng dụng trong công nghệ sinh học và khoa học sự sống.


Giải Cầu lông Sinh viên Trường Đại học Công nghệ – Năm học 2025 – 2026 🔥 Một trong những hoạt động mở màn sôi động của Hội thao UET 2025!



Nhà sáng lập FoodMap (FoodMap.Asia) đang đặt từng bước chân vững chắc trên con đường xây dựng nông nghiệp bền vững. Mới đây, nền tảng thương mại điện tử nông sản FoodMap công bố gọi vốn thành công 2,9 triệu USD vòng pre-series A để mở rộng quy mô hoạt động.

Sâm Ngọc Linh là một trong những cây dược liệu quý hiếm và đặc hữu ở Việt Nam. Nhờ chứa thành phần tự nhiên quý saponin, sâm Ngọc Linh có tác dụng dược lý rất quan trọng, giúp tăng cường hệ miễn dịch và ngăn ngừa ung thư. Vì vậy, loài cây này đã trở thành đối tượng bị sử dụng và khai thác tràn lan, dẫn đến số lượng cá thể tự nhiên giảm sút nghiêm trọng, ảnh hưởng đến nguồn gen quý, đồng thời tạo áp lực cho ngành dược liệu nước ta hiện nay. Đứng trước thực trạng đó, các nhà khoa học trong nước và quốc tế đã sử dụng phương pháp truyền thống kết hợp với kỹ thuật hiện đại nhằm thu thập, bảo tồn và phát triển sâm Ngọc Linh. Trên cơ sở các thành tựu chính đã đạt được trong nghiên cứu về sâm Ngọc Linh hiện nay, các tác giả đã đề xuất một số giải pháp phát triển và nâng cao giá trị thương hiệu quốc gia của loài sâm quý hiếm này.
.jpg)
Ngày 16.10.2020 Khoa Công nghệ nông nghiệp tổ chức buổi giao lưu với các tân sinh viên trên tinh thần đổi mới sáng tạo, truyền cảm hứng và kết nối.

Hiệp hội ra đời là sân chơi chung nhằm tập hợp nguồn lực để thực hiện những nhiệm vụ thực tiễn như: hoàn thiện hạ tầng công nghệ cho nông nghiệp, kết nối quốc tế và mở rộng thị trường.

Ở Việt Nam, lịch sử canh tác hữu cơ đã hình thành từ lâu, nông nghiệp Việt Nam là ngành có lợi thế bởi tính đa dạng và quy mô sản lượng, nhiều loại nông sản có sản lượng hàng đầu thế giới, có nhiều loại nông sản có tiềm năng nhưng chưa được khai thác hợp lý, do đó diện tích nông nghiệp hữu cơ còn khiêm tốn so với tiềm năng. Nông nghiệp hữu cơ là xu hướng tất yếu, song cách tiếp cận như thế nào ở Việt Nam là một vấn đề đáng quan tâm hiện nay. Bài viết phân tích một số thành tựu cũng như những khó khăn trong quá trình ứng dụng sản xuất theo hướng phát triển nền nông nghiệp hữu cơ, trong sản xuất nông nghiệp tại Việt Nam.

Chương trình đào tạo các kỹ sư công nghệ với các chuyên môn: công nghệ thông tin, công nghệ số, điện tử viễn thông, điều khiển, tự động hóa, công nghệ viễn thám, công nghệ nano, công nghệ sinh học có các hiểu biết cần thiết về các lĩnh vực nông nghiệp – cây trồng, vật nuôi và thủy sản – nhằm phát triển và ứng dụng các công nghệ tiên tiến vào nông nghiệp, góp phần tăng hiệu quả của sản xuất nông nghiệp, tạo ra các sản phẩm nông nghiệp có năng suất cao, giá trị cao, an toàn và có sức cạnh tranh trên thị trường trong nước và quốc tế.
.jpg)
Sâm Ngọc Linh là một trong những cây dược liệu quý hiếm và đặc hữu ở Việt Nam. Nhờ chứa thành phần tự nhiên quý saponin, sâm Ngọc Linh có tác dụng dược lý rất quan trọng, giúp tăng cường hệ miễn dịch và ngăn ngừa ung thư. Vì vậy, loài cây này đã trở thành đối tượng bị sử dụng và khai thác tràn lan, dẫn đến số lượng cá thể tự nhiên giảm sút nghiêm trọng, ảnh hưởng đến nguồn gen quý, đồng thời tạo áp lực cho ngành dược liệu nước ta hiện nay. Đứng trước thực trạng đó, các nhà khoa học trong nước và quốc tế đã sử dụng phương pháp truyền thống kết hợp với kỹ thuật hiện đại nhằm thu thập, bảo tồn và phát triển sâm Ngọc Linh. Trên cơ sở các thành tựu chính đã đạt được trong nghiên cứu về sâm Ngọc Linh hiện nay, các tác giả đã đề xuất một số giải pháp phát triển và nâng cao giá trị thương hiệu quốc gia của loài sâm quý hiếm này.
| Địa chỉ: Phòng 312, nhà G2 - 144, Xuân Thủy, Cầu Giấy, Hà Nội | |
| Số điện thoại: (024)32123709 | |
| Email: fat@vnu.edu.vn | |
| Website: fat.uet.vnu.edu.vn |
Bạn đã có tài khoản? Đăng nhập